
任何个体(人或脊椎动物)受到某种特殊刺激(如电休克、阈上剂量的致痫剂)均可出现惊厥发作。但癫痫病人的惊厥阈低于正常人,以致于对健康人无害的刺激、或在感觉阈下的刺激对癫痫病人也足以诱发发作。进一步的机制很复杂,目前尚不完全了解。1952年Brock首先用微电极技术作了开创性研究,开始了在神经元水平研究癫痫的机制,但在神经元水平很难简单地给癫痫活动一个恰当的定义。一大组神经元同步性发放肯定是癫痫的基础,但在正常神经元亦可发现类似的发放,如在视觉注意时于兔海马区可记录到高频发放及尖波,在猫的新皮层于慢波睡眠中多达85%的神经元参与同步性≤0.3H2慢波的形成。
Jefferys等提出下列标准作为癫痫的定义:①爆发迅速出现,而后迅速恢复至相对正常的活动;②破坏正常脑功能;③涉及某一脑区大部分或几乎全部神经元;④在受侵犯区域内大部分神经元的发放率增加。
1.神经传递 了解癫痫机制应首先了解神经元及突触网络的性质。神经冲动的传递遍布全身神经系统,是一个电化学过程,既可发生于单个神经元之内,又可发生于神经元之间,甚至神经与肌肉或腺体之间。
(1)神经元内的传递 单个神经元内的传递为一电过程,是通过细胞膜的电位变化来完成的。膜电位或膜电压的大小是由神经元内外正、负离子的平衡所决定。神经元膜内外电压差是由神经元膜控制离子进出来维持的。神经元的膜不能通过大的负离子,使之不能进入细胞内。Na 、K 、Ca 和Cl一可以通过膜上选择性通道,形成离子电渗而出入神经元。这些通道的一部分是开放的,离子可自由出入神经元。另一些通道为"闸门性"的,其开关有赖于不同的刺激。受控于膜电位变化的通道称为电压门控性通道。 当通道开启时,离子依靠浓度梯度向神经元内外流动,即从浓度高的区域向浓度低处流动。此外,还有主动转运机制,即Na -K 泵的作用,将Na 转运到神经元外,同时使K 进入细胞内。№ -K 泵的作用需要ATP作为能源。
当神经元处于静息状态,Na -K 泵使神经元内较神经元外维持于负电荷状态,称为极化状态,也称为静息膜电位,约为-70mV。静息膜电位反映神经元外№ 和cl-浓度高,而膜内K 和其他阴离子浓度高。
门控性通道的开放使神经元的膜电位发生变化。如果Na 和(或)Ca 通道开放。Na 、Ca 进入神经元内,使膜电位下降,神经元内电荷为正性或负性程度下降,称为去极化,如果K 和(或)Cl-通道开放,K 外流或(和)Cl-内流,使膜电位增加,是为超极化。神经冲动通过膜电位的一系列变化沿轴突传递,包括静息电位,去极化,复极化和超极化,总称动作电位。
一个刺激使神经元膜的局部产生去极化,导致门控性№ 和K 通道开放。Na 迅速进入神经元内,产生去极化;同时K 外流,门控性Na 通道很快关闭,Na 又通过开放的其他通道依照浓度梯度离开神经元内,当№ 外流趋向平衡时,神经元膜开始复极化。但门控性K 通道关闭慢,所以K 达到平衡的时间长,在神经元膜产生短时间的超极化。最后又恢复至静息电位状态,可以接受下一个刺激。
(2)神经元间的传递 神经冲动从一个神经元传到另一个神经元是化学过程。神经冲动到达轴突末端时,轴突释放神经递质至突触间隙,与下一个神经元树突或胞体上的受体结合。神经递质的释放与突触前神经元电压门控性№ 和Ca 通道的开放有关。
神经递质与突触后膜上的受体结合后引起化学门控性离子通道开放,递质的类型决定开放通道的种类,从而决定突触后神经元的反应性质。兴奋性递质使Na 和Ca 通道开放,导致突触后膜去极化,产生兴奋性突触后电位(EPSP)。抑制性递质开启K 和Cl-通道,使突触后膜超极化产生抑制性突触后电位(n,sP)。一个神经元的反应性决定于某一时间内所接受的EPSP和IPSP的总和。这些信号在细胞体或树突进行整合。如果EPSP的总量超过IPSP,神经元膜将去极化,从而对下一个刺激或冲动可以产生反应。如llXSP占优势,产生超极化,对下一个刺激或冲动无反应。

Copyright © 1998 - familydoctor.com.cn All Rights Reserved 家庭医生在线版权所有
本网站敬告网民:网站资讯仅供参考,身体若有不适,请及时到医院就诊。
经营许可证编号:粤B2-20050069号 网络出版服务许可证:总网出证(粤)字第054号 互联网医疗卫生信息服务许可
互联网药品信息服务资格证 短消息类服务接入代码使用证书 粤ICP备14007806号
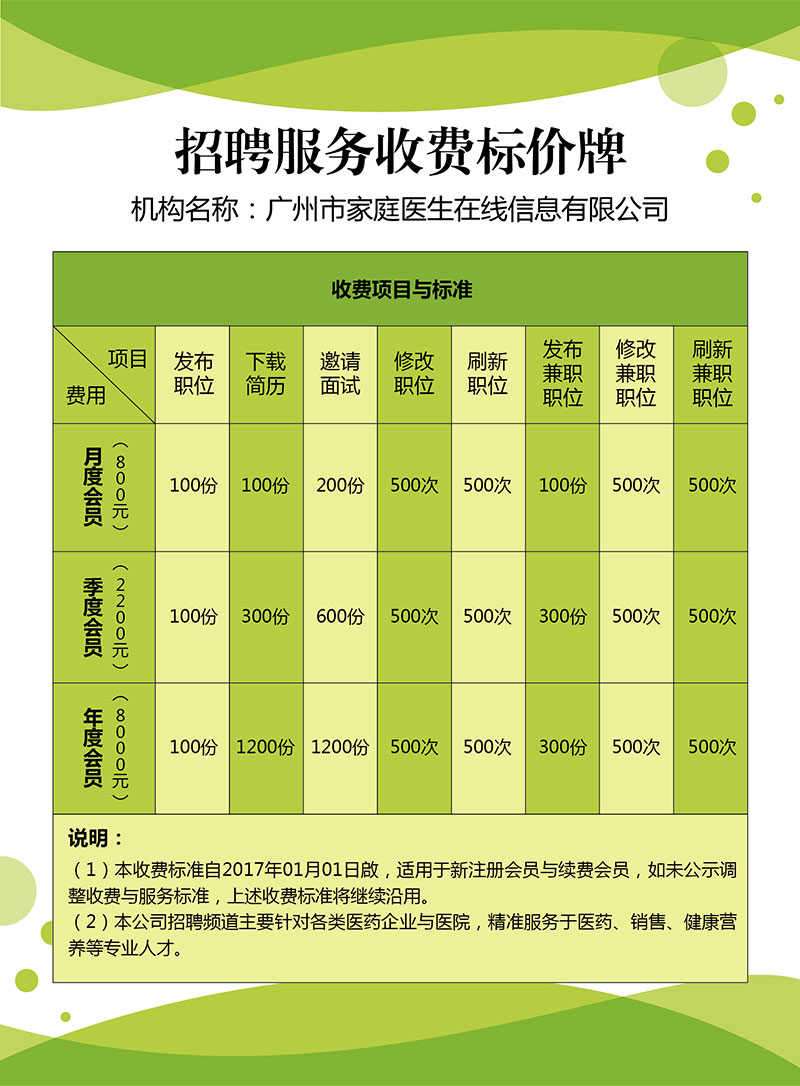
人力资源服务许可证 2019年度广东省“守合同重信用”企业 招聘服务收费标价牌 广播电视节目制作经营许可证 营业执照 食品经营许可证 高新技术企业证书
![]() 粤公网安备 44010402001104号
网络110报警服务
不良信息举报中心
垃圾信息举报中心
广东省通信管理局
粤公网安备 44010402001104号
网络110报警服务
不良信息举报中心
垃圾信息举报中心
广东省通信管理局

![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}